
「内分泌かく乱作用とは」

「詳細編」
両生類の無尾類では、卵からオタマジャクシになり、四肢が伸び、尾が退縮し、やがてカエルになり、陸上生活を営む。また、春先の雨の日に、カエルの大合唱が聞こえてくるように、温度や降雨など季節の環境要因に大きく影響を受けて繁殖活動を行うことも知られている。これらのしくみには、内分泌系(ホルモン)の働きが大きく関わっている。
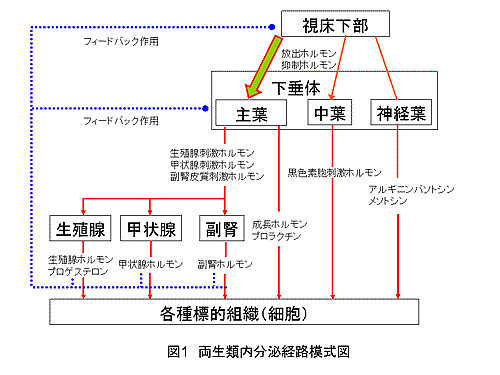
両生類の内分泌系は、他の脊椎動物と同様に、中枢となる「視床下部-下垂体」を軸として、各標的器官のホルモン産生を調節している。図1は代表的な調節経路の模式図を示す。両生類のホルモンの種類や産生場所は、哺乳類と共通する場合が多い。下垂体主葉と中葉から放出されるホルモンは、視床下部から放出される放出ホルモン(生殖腺刺激ホルモン放出ホルモン; GnRH、副腎皮質刺激ホルモン放出ホルモン; CRH、甲状腺刺激ホルモン放出ホルモン; TRHなど)や抑制ホルモン(プロラクチン抑制ホルモン;ドーパミン、成長ホルモン抑制ホルモン;ソマトスタチン)によって調節されている。下垂体主葉からは各内分泌器官に対する刺激ホルモン(生殖腺刺激ホルモン; GTHなど)が放出され、各ホルモン(性 生殖腺ホルモン、甲状腺ホルモンなど)の産生を調節する。これらのホルモンは、血流に乗って体内の各部位に運ばれ作用するとともに、「視床下部-下垂体」へのフィードバック作用も示すことが知られているが、哺乳類に比べて両生類では未解明な点が多い。このほか、抗利尿ホルモンとして知られるバソトシンなどは、視床下部で産生され神経軸索を通して下垂体後葉から放出され、血流を通して標的器官に作用する。
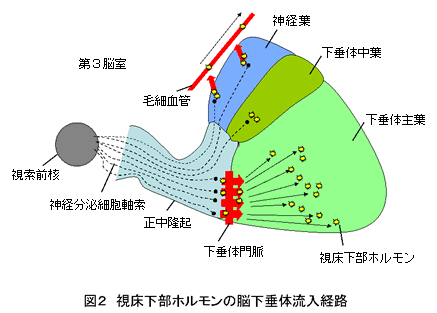
両生類の脳下垂体から分泌されるホルモンは、他の多くの脊椎動物と同様、その産生場所が決まっている。脊椎動物の下垂体の起源は発生学的に同じで、神経下垂体と腺下垂体からなる。図2のように、視床下部にある視索前核の神経分泌細胞から延びる軸索は、下垂体の一部である正中隆起から神経葉にまで達し、神経下垂体ホルモンを分泌する。また、主に下垂体ホルモン放出・抑制を調節するホルモンは軸索を通して正中隆起で分泌され、視床下部と下垂体をつなぐ下垂体門脈を通じて下垂体主葉に流入し、腺下垂体ホルモン産生細胞に作用する(1-3)。下垂体門脈は両生類以上の脊椎動物で発達する。また、下垂体中葉にまで達した軸索から分泌される視床下部ホルモンは、中葉の腺下垂体ホルモン産生細胞に作用する。産生細胞より分泌されたホルモンは、血流に乗って体内の各部位に運ばれ、そこで作用を発現させる。
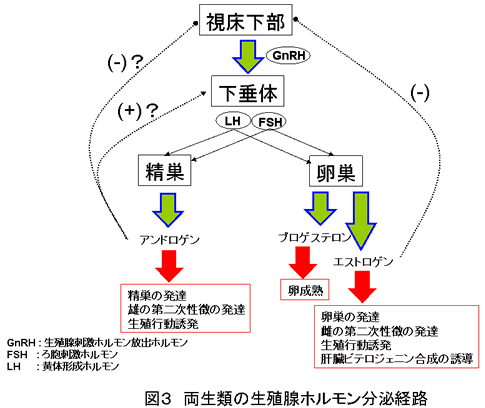
両生類の生殖腺ホルモン分泌経路を図 3 に示す。生殖腺ホルモンの合成は、視床下部から分泌される生殖腺刺激ホルモン放出ホルモン (GnRH) によってコントロールされる( 5 )。両生類の GnRH は、他の多くの脊椎動物と違い、哺乳類型とサケ類型の2種類が存在する。例えば、ウシガエルでは、幼生の頃はサケ型しか存在しないが、成体になるにつれて哺乳類型が増えてくる( 5-6 )。
GnRH の刺激を受け、下垂体から生殖腺刺激ホルモンの分泌が促進される。生殖腺刺激ホルモンには、ろ胞刺激ホルモン( FSH )と黄体形成ホルモン( LH )があり、これらのホルモンは、生殖腺の発達を促進し、生殖腺からのアンドロゲン、エストロゲン、プロゲステロンの産生・分泌を促す( 1, 2, 6 )。ウシガエルでは、精巣からのアンドロゲン分泌は LH により調節される( 6 )。また、卵巣からのエストロゲン分泌は FSH 、プロゲステロン分泌は LH により調節されることが知られている。精巣から分泌されるアンドロゲンは主にテストステロンであるが、両生類では、テストステロンは精巣で 5 α還元酵素によりジヒドロテストステロン( DHT )になって標的器官に作用するといわれている( 5 )。
生殖腺から分泌されたホルモンにおける GnRH に対する抑制機構として、アンドロゲンが視床下部への負のフィードバック、下垂体への正のフィードバック作用を持つといわれており、エストロゲンは視床下部への負のフィードバック作用がある( 5 )。
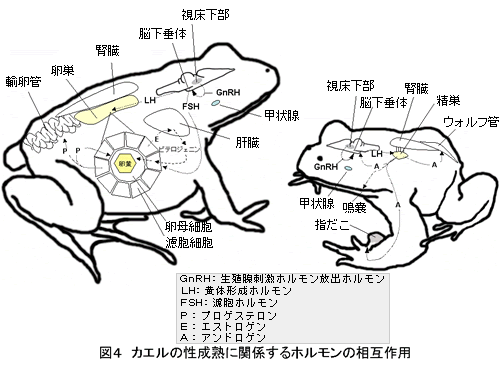
無尾両生類の性成熟に関係するホルモンの流れを図 4 に模式的に示す。性ホルモンであるアンドロゲンとエストロゲンは生殖器官と、第二次性徴の発達をもたらし、生殖行動を誘起させる( 1 ? 2 )。
例えば、雌の卵黄形成において、エストロゲンは、肝臓に作用し、卵黄タンパク質の前駆体であるビテロゲニンの合成を誘導する。ビテロジェニンは血液中を通って濾胞細胞を通して卵母細胞に入るが、卵細胞への取り込みは FSH によって刺激されることがウシガエルで確認されている( 6 )。両生類の場合、一次卵母細胞が十分に成長すると、卵母細胞は繁殖期まで休止状態で保存されるが、繁殖期になるとプロゲステロンが卵母細胞の最終成熟を促進し、排卵へと導く( 1 、 2 )。作用機序は哺乳類とは異なり、生殖腺刺激ホルモンが濾胞組織に作用して、プロゲステロンの産生・分泌を促進し、このプロゲステロンが卵母細胞の受容体に結合し、卵成熟を誘起すると考えられている。また卵管からのジェリー分泌もまた、血中のプロゲステロンの量の変動と相関が認められる。
一方、雄の二次性徴として、雄の指だこや鳴嚢の発達が、一部の無尾両生類において外見的にも顕著に認められる。指だこは、抱接に役立つ器官であり、鳴嚢は繁殖期に同種の雌を呼ぶための breeding call に必要となる。従って二次性徴の発達は繁殖行動のための準備段階といえる。指だこの発達は、精巣の発達や精子形成の進行と密接な時間的関係が見られ( 6 )、精巣から産生されるアンドロゲンの増加によって引き起こされる( 3 )。また、雄の排精は生殖腺刺激ホルモンによって引き起こされることが知られており、この現象を利用して、かつては無尾両生類の排精現象が、妊娠早期診断に用いられていた (6) 。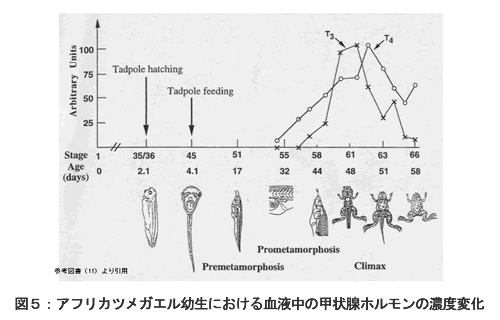
甲状腺ホルモンは、一般的に、発生や成長など、動物の基本的な活動を制御するホルモンであるが、両生類においては、オタマジャクシから成体に変態する過程で、非常に重要な役割を果すホルモンとして良く研究されている。両生類では、甲状腺は左右に一対存在し、濾胞とそれを取り囲む毛細血管の集合体が結合組織に包まれた状態で存在する。甲状腺ホルモンは、甲状腺において分泌されるアミノ酸誘導体のホルモンであり、トリヨードチロニン(T3)とチロキシン(T4)の2種類の化合物が甲状腺ホルモンとして知られている。T3では3個のヨードが、T4では4個のヨードがアミノ酸に付いた構造を持ち、T3はT4よりもはるかに高い生理活性を持つ。各標的器官では、T4がT3へと変換されて作用する。図5は、アフリカツメガエル幼生における血液中の甲状腺ホルモンの濃度変化を示している。両生類の変態期間中には、甲状腺ホルモンの血液中のレベルは著しく上昇し、対象組織中の多くの遺伝子産物が調節を受ける。また、変態上の劇的変化だけでなく、棲性や食性の顕著な変化に連動した生化学的順応をも引き起こす(10)。変態の劇的変化は、甲状腺ホルモンの濃度変化にともなってプログラミングされている。
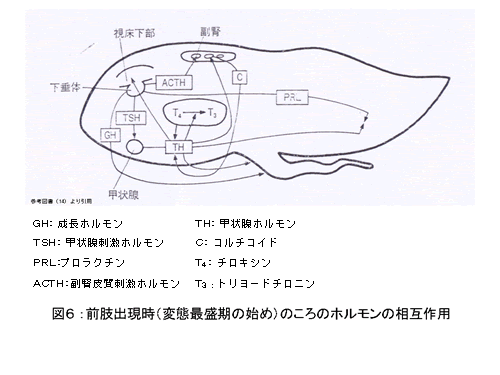
こうした甲状腺ホルモンの血中濃度は、下垂体の前葉から分泌される甲状腺刺激ホルモン( TSH )によって、コントロールされる。前肢出現時(変態最盛期の始め)のホルモンの働きあいの模式図を図 6 に示す。 TSH が、甲状腺において、 T 3 及び T 4 の合成、放出に作用する( 11 )。哺乳類では、 TSH の誘導は、視床下部から分泌される甲状腺刺激ホルモン放出ホルモン( TRH )によって、コントロールされているが、両生類では、 TRH が TSH 分泌を刺激する主要な因子にはなっておらず、 副腎皮質刺激ホルモン放出ホルモン( CRH )が主要な因子である( 13 )。甲状腺から血流に放出され、循環する遊離 T 3 及び T 4 は、 TSH 分泌を抑制的に調節し(負のフィードバック作用)、 CRH の分泌も制御している( 11 )。
副腎から分泌されるコルチコイドもまた、変態作用に関わる。コルチコイドは TSH 同様、副腎皮質刺激ホルモン( ACTH )によって分泌が促進されるホルモンであり、甲状腺ホルモンと相乗効果を持ち、変態を促進させる機能を持つことが知られている( 12 , 14 )。コルチコイドは、変態最盛期の前半に、急激に血液中のレベルが増加する。この時期は、甲状腺ホルモンのレベルが上昇している時期でもあり、甲状腺ホルモンの効果を著しく増強している 。また、腺下垂体主葉で生産・分泌されるプロラクチンは、動物種ごとに多様な生物活性を示すが、両生類の幼生では、甲状腺ホルモンによって促進される変態を抑制する。両生類では、甲状腺刺激ホルモン放出ホルモン( TRH )によって促進的に、ドーパミンによって抑制的にコントロールされる( 3,12,15 )。ウシガエル幼生において、変態最盛期の前にはプロラクチンが変態抑制の方向に働いている。しかし、プロラクチン血中濃度が上昇する変態最盛期後半では、プロラクチンの作用は圧倒的な甲状腺ホルモンとコルチコイドの作用により打ち消され、やがて変態を遂げると考えられているが( 14 )、詳細は未解明である。
参考図書
- 1)
- 岩澤久彰. “本論 1.形態 g. 内分泌器官”. 動物系統分類学 9 (下A1 )脊椎動物(IIa1) 両生類 I. 中山書店, 1996, p. 66-68.
- 2)
- 浦野明央, 石原勝敏 編 “II・3内分泌系”. ヒキガエルの生物学. 裳華房, 1987, p. 98-127.
- 3)
- 日本比較内分泌学会 編. “1. 脊椎動物”. 内分泌器官のアトラス. 講談社, 1987, p. 10-107.
- 4)
- 花岡陽一. “4. 両生類の性分化とホルモン”. ホルモンの生物学9 性分化とホルモン. 学会出版センター, 1984, p. 99-113.
- 5)
- C.A. Herman “3. Endocrinology” Environmental Physiology of the Amphibians. The University of Chicago press, 1992, p. 50-54.
- 6)
- 岩澤久彰, 石田一夫 編. “3 両生類”. 性腺のバイオメカニズム. アイピーシー, 1990, p. 261-357.
- 7)
- 菊山栄・浦野明央. “8. 両生類における生殖行動”. 行動とホルモン. 日本比較内分泌学会編. 学会出版センター, 1983, p. 137-149.
- 8)
- Polzonetti-Mani, A. et al. Growth Hormone and Prolactin in Amphibian Reproduction. Zoological science, vol. 12, 1995, p. 683-694.
- 9)
- Polzonetti-Mani, A. M. et al. Multihormonal control of vitellogenesis in lower vertebrates. International Review of Cytology, vol. 239, 2004, p. 1-46.
- 10)
- 金森章. “8. 甲状腺ホルモンと発生”.ホルモン分子生物学4 甲状腺ホルモン 日本比較内分泌学会編.学会出版センター,1998,p.129-136.
- 11)
- Shi, Y-B. Amphibian metamorphosis from morphology to molecular biology. Wiley-Liss Press, 2000, p. 22-27.
- 12)
- Kikuyama, S. et al. Aspects of amphibian metamorphosis:hormonal control. International Review of Cytology, vol. 145, 1993, p.105-148.
- 13)
- Denver, R.J. The molecular basis of thyroid hormone-dependent central nervous system remodeling during amphibian metamorphosis. Comparative Biochemistory and Physiology Part C 119, 1998, p.219-228.
- 14)
- 菊山栄.“3.両生類の変態とホルモン”.組織分化とホルモン -新しい実験形態学の展開- 新井康充・平野哲也編.学会出版センター,1984,p.39-42.
- 15)
- 小野雅夫. “1. 成長ホルモン・プロラクチン・ソマトラクチン”. ホルモンの分子生物学2 成長ホルモン・プロラクチンファミリー. 日本比較内分泌学会編. 学会出版センター, 1996, p. 1-26.
- 16)
- 菊山栄. “Ⅱ-3. 成長期と変態期-両生類における研究”. ホルモンと時間. 石居進編. 学会出版センター, 1980, p. 43-56.